生化学
沿革
生化学講座は昭和49年4月に慶応義塾大学から着任した間宮群二教授によって開設された。この時点では生化学の講義等は開始されていなかったが、間宮教授は入間基地内に設けられた木造の仮研究室において、昭和51年3月の所沢移転まで静かな環境のもと研究活動を行った。
昭和54年4月にドイツから帰国した後藤完教授を初代教授として生化学第二講座が新設され、旧生化学講座は生化学第一講座と改称された。生化学第一講座では当初はタンパク質の一次構造決定と作用機序の解明を、後には遺伝子工学や細胞培養の技術を取り入れ細胞内情報伝達やがん転移抑制機構の研究を行った。
生化学第二講座では血管構成細胞の生理化学的研究を主要テーマとして掲げ、主に組織培養法による研究を展開した。
平成元年度に後藤教授の退官に伴い、生化学第二講座二代目教授として山本三毅夫教授が九州大学より着任し、改良遺伝子発現解析法による疾患関連遺伝子群のプロファイリングと機能解析を講座のテーマとして精力的に研究を行った。生化学第一講座においては、平成6年4月に間宮教授の後任として瀧嶋邦夫助教授が教授に昇任した。平成20年4月に山本教授の退官に伴い生化学第一講座と第二講座は統合され、再び生化学講座が開設された。
平成31年4月に瀧嶋邦夫教授の退官に伴い佐藤泰司教授が着任し現在に至っている。現在、教授以下今村宰准教授、新井仁明助教、伊達木穣助教の4名が教育および研究活動に従事している。
教育の概要
第1、2学年に対する講義と実習、並びに高等看護学院における生化学の講義を担当している。授業は、内容を生体物質、代謝、遺伝子の3分野に分け、生体物質ではタンパク質、糖、脂質、核酸などの構造とこれら生体分子の機能面にあたる酵素反応や膜輸送などを、代謝では糖代謝、脂質代謝、アミノ酸代謝、ヌクレオチド代謝および代謝の全体像を、遺伝子では遺伝現象の基本である複製、修復、転写、翻訳、遺伝子発現調節について講義と演習を行っている。
また、病態を生化学・分子生物学の観点から理解することを期待して種々の疾患を題材とした学生発表も行っている。これらを通して生命現象を分子レベルで理解し、臨床医学の現場で生化学の知識と考え方を活用できる人材が育成されることを期待している。講義と並行して年間13回行われる実習も重視し、講義内容のより深い理解につながるような実験テーマを設けている。
具体的には、グリコーゲンの精製と定量、ヘモグロビンの調製とサブユニット構造の解析、赤血球を用いた生体膜の性状解析、肝臓アルカリホスファターゼの精製と酵素反応実験、ヒトアセトアルデヒド脱水素酵素の遺伝子多型分析とタンパク質発現解析、DNA二重鎖切断やチミンダイマー形成によるDNA損傷の大腸菌を用いた解析などである。
これらの実験では、マイクロプレートリーダーを用いたタンパク質定量、SDSポリアクリルアミドゲル電気泳動法とウェスタンブロッティング法、PCR法、基本的な遺伝子操作法、ラクトースオペロンを用いた遺伝子発現誘導などの実験技法に親しめるよう工夫している。
研究の要約
当講座では、現在、神経疾患の治療を念頭に置いて主に神経の細胞分化と細胞死に焦点をあてた研究を行っている。これまで、MAPキナーゼファミリーに属するERK2の神経細胞分化と神経機能における役割を解明するため、マウスErk2の遺伝子構造を決定し、この結果に基づいてErk2遺伝子ターゲッティングマウスを理化学研究所との共同研究で作出している。Erk2ノックダウンマウスの解析からERK2は空間学習や恐怖条件付け学習などにおける記憶・学習過程に重要な役割をもつことを明らかにした。
そして、条件付きErk2ノックアウトマウスの神経幹細胞を解析することにより、ERK2は神経幹細胞の自己複製に必須であり、グリア細胞への分化誘導を抑制することを見出した。さらに、Erk1/2ダブルノックアウトマウスの解析により、大脳皮質層構造の構築における神経細胞の移動にもERK1/2が関与していることを明らかにしている。
また、てんかんモデルラットの脳組織で発現が増加している機能未知遺伝子が遺伝子発現プロファイリング解析によって複数個得られており、その中の1つが挫傷脳組織において発現が増強されていることを見出した。この遺伝子を培養細胞で強制発現させるとアポトーシスが誘導され、この遺伝子に対するsiRNAを培養細胞に導入するとグルタミン酸やCaイオンの濃度上昇で引き起こされる細胞死が抑制されることを明らかにした。技術開発的な研究としては、内在性あるいは移植神経幹細胞を効率よく神経細胞に分化誘導する薬剤のスクリーニング系を開発し、臨床薬剤の中から新たな作用を有するものを複数同定することに成功している。また、局所限定的な遺伝子発現を非侵襲的に操作する系の開発も進めている。

遺伝子改変マウス
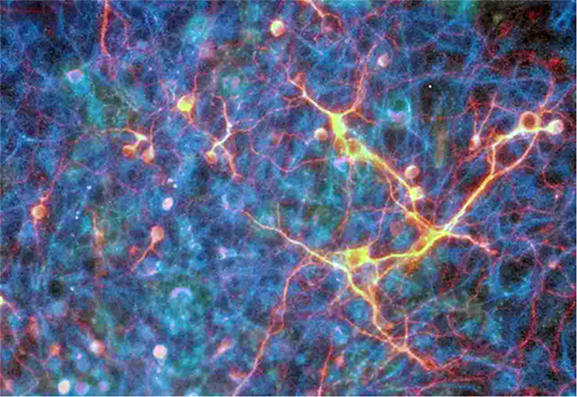
神経幹細胞の分化誘導





